Материалы этого раздела курса направлены на понимание основных эффектов на
клеточном, тканевом, организменном, популяционном уровнях при действии на
человека ионизирующих излучений на Земле и в условиях космического пространства.
Они знакомят с методами количественных оценок ближайших и отдаленных
неблагоприятных последствий действия радиации в зависимости от дозы и мощности
дозы радиационного воздействия.
Рассматриваются вопросы биологической эффективности различных видов излучений и
особенности действия на организм Галактических космических лучей (ГКЛ),
солнечных космических лучей (СКЛ) и протонов радиационных поясов Земли.
Представляются подходы к определению суммарного радиационного риска для
космонавтов в течение всей их жизни и оценки возможного сокращения ее
продолжительности в результате осуществления межпланетного полета к Марсу и
длительных полетов на орбитальных станциях «Мир» и «МКС».
Эти материалы обобщают результаты экспериментальных и теоретических исследований
по проблеме обеспечения радиационной безопасности космических полетов,
проводимых с 1964 г в Институте медико-биологических проблем МЗ СССР, Институтах
биофизики АН СССР и Минздрава СССР, Государственном НИИИ Военной медицины МО
СССР и др.
Некоторые основы биологического действия источников излучений с различной
плотностью ионизации на живые организмы
Ионизирующие излучения
Ионизирующие излучения – один из основных экологических факторов, который
постоянно действовал в процессе эволюции на окружающую среду, в том числе на
живые организмы на Земле, и в околоземном пространстве. Этот фактор необходимо
изучать максимально всесторонне и тщательно как в плане исследования
неблагоприятного воздействия на окружающую среду и человека, так и в отношении
возможности полезного использования его в промышленности и в медицине для
сохранения здоровья человека.
Исследования по радиобиологии проводятся уже в течение 110 лет с момента
открытия рентгеновских лучей и явлений радиоактивности. Исключительно
разнообразен набор объектов, являющихся предметом радиобиологических
исследований. Это – макромолекулы, фаги, вирусы, простейшие, клеточные, тканевые
и органные культуры, многоклеточные растительные и животные организмы, человек,
популяции, биоценозы.
Радиобиологические исследования всегда проводились при строгой количественной
оценке поглощенных доз в клетках и в различных тканях организма. Единицей
поглощенной дозы является 1 Гр = 1 Дж/кг.
Термины
Дозиметрические единицы и их определение в
СИ и СГС
СИ
СГС
Экспозиционная доза
За единицу экспозиционной дозы принят 1 кулон на килограмм (Кл/кг), когда
сопряженная корпускулярная эмиссия производит в 1 кг сухого атмосферного
воздуха ионы, несущие один кулон электричества каждого знака
Рентген –
единица экспозиционной дозы рентгеновского или гамма-излучений, при
прохождении которого образующаяся корпускулярная эмиссия в 1 см3 сухого
атмосферного воздуха при 0о С
и давлении 760 мм рт. ст. после завершения всех ионизационных процессов
создает ионы, несущие 1 электростатическую единицу количества
электричества каждого знака ( 2,08·109 пар ионов). 1 рентген = 1 ед.СГС/см3
(1
р = 2,58•10-4 Кл/кг).
Поглощенная доза D=de/dm
Величина энергии ионизирующего излучения, переданная веществу.
Единицей является 1
Грей – энергия в 1
Джоуль, переданная массе в 1 кг 1Гр = 1 Дж/кг = 100 рад
Рад -
единица поглощенной дозы, когда грамму вещества передается энергия 100
эрг. 1 рад = 100 эрг/г = 10-2 Гр
= 1 сГр
При экспозиционной дозе 1 р поглощенная доза в ткани с точностью до 10%
численно равна 1 раду
Эквивалентная доза (HT,R)
Поглощенная доза в органе или ткани DT,R, умноженная
на соответствующий взвешивающий коэффициент для данного вида излучения
WR (называемый
также коэффициентом качества излучения (КК)).
HT,R = WR • DT,R
Единицей является 1 Зиверт 1 Зв = 1 Гр • WR = 100 бэр
Единицей эквивалентной дозы являлся
1
бэр 1 бэр=1 рад • КК 1 бэр = 10-2Зв = 1 сЗв
Одним из основных парадоксов радиобиологии, постоянно
занимавшим умы исследователей на ранних этапах исследований, является
несоответствие между ничтожной величиной поглощенной энергии и крайней степенью
выраженности реакций биологического объекта вплоть до летального эффекта.
Несмотря на существующие в природе колоссальные различия в
чувствительности к ионизирующим излучениям различных биологических видов,
облучение в дозе 10 Гр убивает всех млекопитающих. В то же время в биологической
ткани в объеме 1 мкм3,
содержащем 1010 атомов,
создается лишь около 200 ионизаций (примерно такое же число реагирующих
молекул). Другими словами, смертельная доза в 10 Гр (10 Дж/кг) способна изменить
энергетический статус ничтожно малого числа молекул в данном объеме. При
облучении человека массой 70 кг в этой дозе поглощается всего 700 Джоулей (170
калорий). При этом максимальный нагрев тела не превысит 0,001о С
(тепловой эффект эквивалентен примерно стакану выпитого горячего чая).
С целью объяснить радиобиологический парадокс, связанный с
несоответствием между ничтожно малым количеством поглощенной энергии излучения и
возникающими экстремальными эффектами, приводящими к гибели большинства
биологических объектов, в 1922 г. Ф. Дессауэр предложил первую теорию,
объясняющую радиобиологические эффекты повреждения клеток различных тканей
дискретностью событий – актов ионизации в чувствительном объеме клетки.
Эти представления получили в последующем свое развитие в виде
принципа попадания и теории мишеней в работах Н.В.
Тимофеева-Ресовского, К
Циммера, Д. Ли и ряда других исследователей.
После открытия повреждающего действия ионизирующих излучений (ИИ),
выражающегося в гибели клеток различных тканей, а также гибели животных,
растений и других биологических видов, было обнаружено, что величины доз,
приводящие к летальным эффектам, различаются в широких пределах, порой на
несколько порядков. Другими словами, каждому биологическому объекту (различным
клеткам, тканям, органам и целым организмам) свойственна своя мера
восприимчивости к воздействию ионизирующей радиации – видовая
радиочувствительность.
Радиочувствительность может сильно варьироваться и в пределах одного вида и
характеризуется понятием "индивидуальная радиочувствительность".
В соответствии со сформулированным еще в 1903 г.
французскими исследователями И.
Бергонье и Л.
Трибондо законом степень
поражения делящихся клеток в размножающихся клеточных популяциях тем выше, чем
большая у них способность к размножению, чем выше скорость их размножения. В
процессе последующей трансформации клеток становится более выраженной их
функциональная специализация, и эти клетки становятся менее
радиочувствительными. Этот закон является справедливым до настоящего времени.
Наряду с радиочувствительными быстро обновляющимися тканями
(кроветворная система, эпителий слизистой тонкого кишечника, эпидермис) имеется
целый ряд радиорезистентных (радиоустойчивых) тканей. Это мышечная, костная,
нервная ткани, сосудистый эпителий и др..
Одной из наиболее радиочувствительных тканей, определяющей
устойчивость (способность выживания организма) в ближайшем периоде после
равномерных облучений млекопитающих является система кроветворения. Продукция
кроветворных клеток осуществляется непрерывно в течение жизни за счет
пролиферации (деления) стволовых кроветворных клеток и их дифференцировки
(трансформации) в костном мозге в морфологически идентифицируемые
предшественники нейтрофилов, лимфоцитов, тромбоцитов и эритроцитов.
У человека в час обновляется около (0,3-0,7) 109 кроветворных
клеток на 1 кг массы тела. Повреждение стволовых кроветворных клеток приводит к
уменьшению числа способных к делению костномозговых кроветворных клеток и
уменьшению концентрации функциональных клеток в периферической крови.
Уменьшение числа клеток белого ряда нейтрофилов и лимфоцитов
приводит к снижению иммунитета организма и его способности сопротивления
болезнетворным бактериям и вирусам и способствует развитию инфекционного
процесса.
Значительное снижение числа тромбоцитов нарушает процесс
нормальной свертываемости крови и увеличивает вероятность кровоизлияния в
различные органы и ткани.
Снижение числа эритроцитов и гемоглобина нарушает дыхательную
функцию по доставке кислорода к тканям. Все это приводит к снижению
жизнеспособности и увеличению вероятности гибели организма в ближайшем периоде
после острых облучений в летальном диапазоне доз.
В таблице 1 представлены значения среднелетальных доз
ЛД 50/30-60 для различных биологических объектов.
Таблица 1
Значения среднелетальных доз ЛД 50/30-60 для
биологических объектов, сГр
Биологический вид
ЛД 50/30-60
Биологический вид
ЛД 50/30-60
Мышь, крыса
850-940 / 800-900
Птицы
800-2000
Хомяк
900-940 / 850-900
Рыбы
800-2000
Кролик
/ 840-890
Змеи
8000-20000
Морская свинка
/ 255
Насекомые
1000-10000
Овца
235-250 / 155
Растения
1000-15000
Коза
325-330 / 230
Бактерии Micrococcus radiodurens
105-106
Свинья
400 / 195
Собака
320-330 / 255-260
Обезьяна
550-620 / 400
Человек
350-400 / 225-350
Примечание: первые значения для млекопитающих соответствуют
максимальной поверхностной поглощенной дозе, вторые значения − среднетканевой дозе
Из данных таблицы видно, что среднелетальные дозы для крупных млекопитающих
оказались в 2,5-3 раза более низкими, чем для мелких лабораторных животных,
свидетельствуя о большей их радиочувствительности. Наиболее устойчивыми являются
змеи, насекомые, растения и особенно бактерии. Наиболее обширное изучение действия радиации на различные ткани началось после
возможности подробного исследования структур и функций различных органелл клеток
с помощью электронных микроскопов, изучения процессов клеточного деления и
передачи наследственной информации от материнских клеток к дочерним. Особое
значение для понимания механизмов принадлежит исследованиям нуклеиновых кислот,
которые были открыты еще в 1870 г., но огромная биологическая роль которых стала
ясна лишь спустя 50 лет. С ними оказался связан процесс передачи наследственной
информации, стали понятными процессы синтеза белков. Было выяснено, что наследственная информация дискретна, ее составляют
многочисленные гены, расположенные вдоль хромосом в линейном порядке. Каждый ген
занимает постоянное определенное место (локус) в определенной хромосоме.
Генетическая информация, необходимая для развития целого организма, содержится
только в полном гаплоидном наборе хромосом. Стало понятным наличие особых наиболее чувствительных элементов клеток
(мишеней), нарушение структуры и функции которых при минимальных выделениях
энергии может нарушить основной принцип функционирования размножающихся тканей,
связанный с делением клеток и передачей наследственной информации от материнских
к дочерним клеткам.
Нуклеиновые кислоты
Постоянство числа, индивидуальность и сложность строения,
авторепродукция и непрерывность в последовательных генерациях
клеток размножающихся тканей говорят о большой биологической
роли нуклеиновых кислот.
В состав нуклеиновых кислот входят углерод, кислород, азот и
фосфор. Известны две группы кислот: рибонуклеиновая кислота (РНК)
и дезоксирибонуклеиновая кислота (ДНК). Они отличаются
химическим строением и свойствами. Согласно последним данным,
ДНК находится не только в ядре, но входит также в состав
органоидов цитоплазмы, например митохондрий.
Основные хранители РНК, ядрышки, находящиеся в ядре, и рибосомы,
в которых происходит синтез белков. Кроме того, РНК находится в
цитоплазматическом матриксе - цитоплазме.
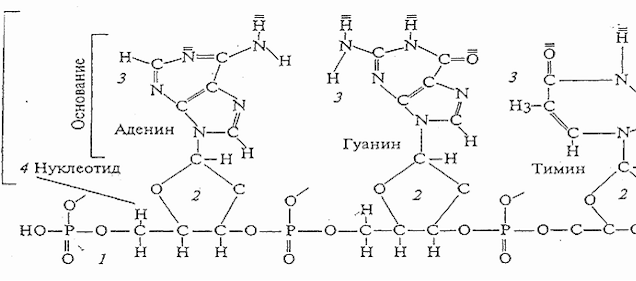
Нуклеиновые кислоты представляют собой биополимеры, мономерами которых служат
нуклеотиды. В каждый нуклеотид входит молекула рибозы в РНК или дезоксирибозы в
ДНК, связанные с молекулой фосфорной кислоты, и одно из 4-х азотистых оснований: аденин
(А), гуанин (Г), цитозин (Ц) и тимин (Т) (или урацил в составе РНК).Простейшие
нуклеиновые кислоты – мононуклеотиды, т.е. отдельные нуклеотиды. Отдельные
нуклеотиды при полимеризации, будучи соединенными между собой остатками
фосфорной кислоты, образуют полинуклеотиды, молекулы которых могут быть
образованы десятками, сотнями и тысячами нуклеотидов. Нуклеотиды могут
располагаться в различной последовательности, что обеспечивает большое
разнообразие соединений этого класса. (Рис. 1).
Основная биологическая функция ДНК заключается в хранении, постоянном
самовозобновлении, самовоспроизведении (репликации) и передаче генетической
(наследственной) информации в клетке. Биологическая роль РНК связана
преимущественно с синтезом белка, т.е. с реализацией наследственной информации.
Именно РНК является посредником между ДНК и строящимися в рибосомах белковыми
молекулами.
Рис. 1 Строение одной из двух нитей ДНК:
1- сахарофосфатный скелет;
2- дезоксирибоза;
3- азотистые основания аденин, гуанин, тимин;
4 - нуклеотид.
Молекула ДНК состоит из двух спирально закрученных нитей. Азотистое
основание одной из нитей ДНК связано водородным «мостиком» с
основанием другой нити, причем так, что аденин может быть связан
только с тимином, а цитозин – только с гуанином. Они комплементарны
(дополнительны) друг другу. Порядок расположения оснований в одной
цепи определяет порядок в другой. Именно на этом основано особое
свойство: кодирования информации о большом многообразии необходимых
белков и способность к самовоспроизведению, т.е. к автопродукции.
В настоящее время известно более 20 аминокислот, из которых 20
входят в состав белковых молекул. В то же время молекулы многих
белков содержат более 100 мономеров (различных аминокислот), которые
соединены одна за другой, подобно бусам на нитке. Перестановка или
замена всего одной-единственной аминокислоты влечет за собой
значительные изменения свойств. Так, например, в молекуле
гемоглобина около 300 мономеров (аминокислот), но при замене одной
из них – глутаминовой кислоты – валином свойство гемоглобина в
отношении передачи кислорода тканям резко снижено. Люди с таким
аномальным гемоглобином страдают наследственным заболеванием -
серповидноклеточной анемией.
В структуре ДНК отражена последовательность расположения
триплетов азотистых оснований (кодонов), которая определяет
генетическую информацию клетки. При этом участок ДНК, содержащий
информацию о структуре какого-либо одного белка принято называть
геном.
Четыре азотистых основания, входящие в состав молекулы ДНК, в
комбинациях по три дают 64 разных кодона. Этого более чем достаточно
для кодирования 20 аминокислот. Эта информация считывается в
процессе транскрипции информационной РНК (иРНК). Она определяет
порядок расположения аминокислот в белках при последующем их синтезе
в рибосомах.
В процессе репликативного синтеза и удвоения ДНК структура ДНК
точно воспроизводится, что позволяет произвести позже точное и
равное распределение генетического материала между материнской и
дочерней клеткой в процессе ее деления.
Молодые клетки, образовавшиеся после деления, не могут сразу
немедленно приступить к новому клеточному делению. В них прежде
должны произойти важные процессы:увеличение объема,
восстановление структурных компонентов ядра и цитоплазмы,
связанных с синтезом нуклеиновых кислот и белка.
Совокупность процессов, происходящих в клетке от одного деления
до следующего и заканчивающихся образованием двух клеток новой
генерации, называется митотическим циклом. Общая длительность
клеточного цикла для кроветворных клеток костного мозга в норме
составляет около 12-24 часов, у крипт кишечного эпителия от 10
до 20 часов. У ряда клеток других тканей он может составлять
несколько суток или даже недель. Различают 4 периода этого
цикла: пресинтетический – G1, синтетический – S,
постсинтетический – G2 и митоз (М).
В нормальных условиях при отсутствии воздействия ионизирующих
излучений (ИИ) под действием высокореактивных химических
радикалов, а также физических агентов незначительной
интенсивности могут иметь место нарушения структуры ДНК. Эти
нарушения обычно быстро восстанавливаются в клетке под действием
ферментов репарации.
В клеточном цикле имеется несколько так называемых сверочных
точек, «чекпойнтов» (в конце стадии G1 и в начале G2), при
прохождении которых ферментативные системы проверяют ДНК на
повреждения, и в случае их выявления активируют репарационные
процессы и происходит небольшая задержка продвижения клетки по
циклу с целью восстановления структуры ДНК и нормального
завершения процесса репликации и удвоения ДНК до вступления
клетки в митоз.
При воздействии ИИ имеют место уже значительные повреждения и та же система
приводит к задержке начала синтеза ДНК, увеличению по длительности S периода
и удлинению периода подготовки клетки к делению G2. Блок в прохождении
клеток по циклу нагляднее всего проявляется в виде зависимой от дозы
задержки наступления первого постлучевого митоза.
При увеличении дозы системы репарации не могут обеспечить необходимый
уровень восстановления клетки. Образуются хромосомные и хроматидные
нарушения структуры (хромосомные аберрации), образуются дицентрики,
ацентрические фрагменты хромосом, мосты, межхромосомные обмены и др., и не
происходит нормального равного распределения хромосом между дочерними
клетками.
Аберрации, сопровождающиеся образованием ацентрических фрагментов, мостов и
дицентриков, получили название нестабильных, так как приводят к гибели либо
самой клетки, либо ее ближайших потомков.
Перестройки, сопровождающиеся только перемещением участков поврежденных
хромосом, когда весь генетический материал остается связанным с центромерой
и может равномерно распределяться между дочерними клетками, относят к
стабильным, так как они могут передавать генетический материал из поколение
в поколение, сохраняясь в организме в течение многих лет. Примером могут
служить транслокации, когда участок генома перемещается в новое для него
место в молекуле ДНК, но продолжает функционировать. Это
может приводить к воспроизводству измененных белков, появлению различных
заболеваний, в том числе к увеличению вероятности развития опухолей.
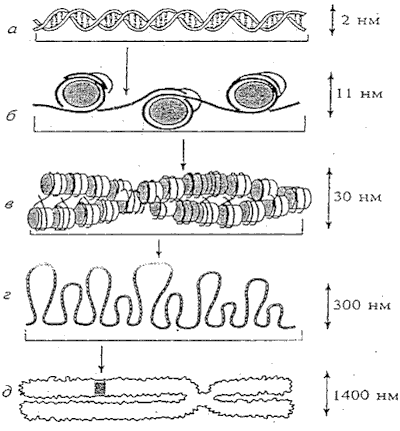
Рис.2. Строение участка молекулы ДНК и ее пространственная упаковка в процессе
последовательной спирализации после завершения синтеза ДНК и подготовки клетки к
делению. Последний фрагмент рисунка - хромосома на стадии метафазы в митозе до
разделения.
При облучении в клетке повреждаются многие ее структуры. Из-за большого размера
и особо важной роли молекул ДНК в передаче наследственной информации в качестве
основной мишени наиболее часто рассматривается характер возникающих нарушений
ДНК и возможность их репарации (восстановления). В клетке различные участки ДНК
одной и той же молекулы находятся очень близко друг к другу из-за многократного
сворачивания нитей ДНК в структуры все большего диаметра при подготовки клетки к
делению (Рис.2).
ДНК, РНК − Возможности их репарации
Постоянство числа, индивидуальность и сложность строения,
авторепродукция и непрерывность в последовательных генерациях
клеток размножающихся тканей говорят о большой биологической
роли нуклеиновых кислот.
В состав нуклеиновых кислот входят углерод, кислород, азот и
фосфор. Известны две группы кислот: рибонуклеиновая кислота (РНК)
и дезоксирибонуклеиновая кислота (ДНК). Они отличаются
химическим строением и свойствами. Согласно последним данным ДНК
находится не только в ядре, но входит также в состав органоидов
цитоплазмы, например, митохондрий.
Основные хранители РНК, ядрышки, находящиеся в ядре, и рибосомы,
в которых происходит синтез белков. Кроме того, РНК находится в
цитоплазматическом матриксе - цитоплазме.
В отличии от редкоионизирующих излучений при облучении в той же
дозе от источников радиации с высокими линейными потерями
энергии (ЛПЭ), с высокой плотностью ионизации и выделении в
рассматриваемом объеме клетки локально большого количества
энергии (галактические космические лучи, ускоренные в
эксперименте многозарядные ионы (УМИ), быстрые нейтроны)
вероятность множественных двойных разрывов резко увеличивается и
снижается возможность их репарации.
Это приводит к появлению множественных видов хромосомных
аберраций при митозе и значительному увеличению вероятности
гибели клеток. Все это обусловливает существенно большую
относительную биологическую эффективность (ОБЭ) излучений с
высокими значениями ЛПЭ в отношении поражения клеток различных
тканей.
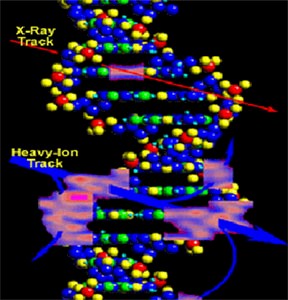
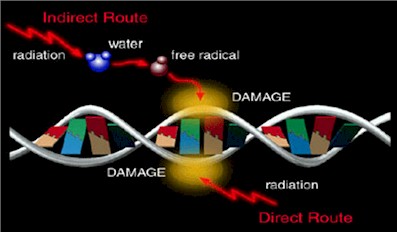
Радиационные повреждения молекулы ДНК
На схеме отчетливо видно более глубокое радиационное повреждение
молекулы ДНК при действии ускоренных тяжелых ионов с высокими
линейными передачами энергии (ЛПЭ) по сравнению с воздействием
рентгеновского излучения.
Ионизирующие излучения, действуя на водную среду, в которой
находятся клетки, образуют целый ряд высокоактивных химических
радикалов, в свою очередь, способных вызывать серьезные структурные
повреждения ДНК и других клеточных органелл, нарушая их нормальную
функцию (так называемое косвенное действие ионизирующих излучений).
Ниже представлена схема образования высокореактивных радикалов,
продуктов радиолиза воды и последующих радикалов, связанных с
взаимодействием с растворенными в воде органическими соединениями.
Следует отметить, что если прямое повреждающее действие
осуществляется, можно считать, практически мгновенно, то
опосредованное действие носит существенно более длительный характер
с образованием порой токсичных белковых продуктов. При этом прямое
непосредственное действие ответственно лишь за 10-20 % лучевого
поражения.
На схеме наряду с прямыми повреждениями молекулы ДНК за счет
ионизации показано непрямое (косвенное) поражение структуры ДНК за
счет образования под действием радиации высокореактивных свободных
радикалов.
Косвенное действие, при котором поражение критических структур
осуществляется продуктами радиолиза воды, может достигать и быть
ответственно за 80-90 % лучевого поражения, которое действует
дистанционно, как в пространстве, так и во времени.
Множество
эффектов непрямого действия ионизирующих излучений (ИИ)
зафиксировано по эффектам в полностью экранированных тканях
организма при облучении соседних и даже удаленных участков тела.
Наиболее часто именно косвенное действие радиации усиливается или
ослабляется с помощью химических модификаторов.
Кривые
выживаемости клеток почки человека при действии излучений с
различными ЛПЭ
Рис.3. Кривые выживаемости клеток почки человека при действии
излучений с различными ЛПЭ
а) рентгеновское излучение 250 кВ,
ЛПЭ =1,3 кэВ/мкм;
б) нейтроны с энергией
15 МэВ, ЛПЭ = 80 кэВ/мкм;
в) α частицы 210Ро,
ЛПЭ =166 кэВ/мкм.
Светлые значки – облучение в присутствии кислорода, темные –
облучение в условиях аноксии.
Особенно сильным модификатором, усиливающим повреждающее действие
ИИ, является кислород. Радиобиологические исследования по облучению
клеток различных тканей в условиях присутствия в них кислорода и при
аноксии (отсутствии кислорода) показали, что равноэффективные дозы в
первом случае оказываются приблизительно в 3 раза меньшими (так
называемый коэффициент кислородного усиления (ККУ) радиационного
поражения).
Большее повреждающее действие ИИ в присутствии кислорода в
клетках связано, с одной стороны, с тем, что усиливается
концентрация высокореактивных радикалов и особенно перекисных
соединений, что усиливает структурные повреждения ДНК и других
структур клетки, а с другой - при ионизации атомов на одном из
участков макромолекулы образуется неспаренный электрон, который
захватывается кислородом на свою орбиту, и кислород присоединяется к
молекуле ДНК в месте разрыва одной из химических связей. Такая
модификация снижает эффективность ее репарации.
На Рис.3 представлены кривые выживаемости клеток некоторых тканей,
находящихся в кислородной среде и в условиях аноксии, в зависимости
от поглощенной дозы при воздействии редкоионизирующих и
плотноионизирующих излучений. Можно отметить больший повреждающий
эффект и более выраженное снижение доли выживших клеток при
облучении в присутствии кислорода, что особенно выражено при
воздействии редкоионизирующих излучений (ККУ равен от 2,5 до 3). При воздействии плотноионизирующих излучений на клетки
тканей, находящихся в кислородной среде, имеет место незначительное
увеличение радиационного поражения клеток и даже отмечается
отсутствие кислородного эффекта.
Выживаемость клеток радиочувствительных тканей организма при действии
излучений с различными ЛПЭ в зависимости от поглощенной дозы
Выживаемость клеток различных тканей в зависимости от дозы
облучения широко начала изучаться с 1956 г. после разработанного
метода культивирования клеток вне организма (in vitro) в
термостате в чашках с питательной средой. Долю выживших клеток
после облучения в различных дозах определяли на основе отношения
числа нормальных полноценных колоний клеток, выросших в
облученных и контрольных чашках в результате многократного их
деления.
Эта идея позднее была применена к тканям in vivo (в живом
организме). Канадские ученые Дж. Тилл и Е. Мак-Кулох в 1961 г.
предложили метод определения радиочувствительности стволовых
клеток гемопоэтической ткани. Они ввели клетки костного мозга в
хвостовую вену летально облученным мышам-реципиентам, у которых
через 7-9 дней на селезенке появились видимые невооруженным
глазом колонии, состоящие из потомков введенных клеток.
Путем сравнения числа колоний, образующихся при введении клеток
костного мозга от облученных в разных дозах и необлученных
мышей, были получены типичные кривые выживаемости стволовых
кроветворных клеток. Этот метод, получивший название
экзоколониального теста, широко используется и в настоящее время
для оценки радиочувствительности стволовых кроветворных клеток
донора.
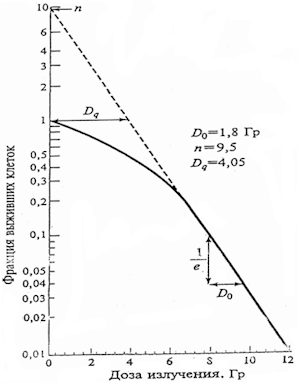
Известно, что процесс ионизации структур клетки носит
стохастический характер. Для инактивации клетки при действии
плотноионизирующих излучений, как правило, бывает достаточно
реализации одного события попадания частицы в ядро клетки и
повреждения структуры ДНК. Кривые выживаемости клеток различных
тканей в этом случае описываются экспоненциальной зависимостью
от дозы:
N = N0exp(−D/D0),
где D0 − параметр, характеризующий радиочувствительность ткани,
- дозу, при которой доля выживших клеток, избежавших
взаимодействия, будет снижена в е раз.
При действии редкоионизирующих излучений, учитывая активную и
достаточно оперативную работу репарационных ферментативных
систем в клетках, для реального закрепления радиационного
поражения в клетке, чтобы она потеряла свой пролиферативный
потенциал (способность осуществления большого числа делений и
образованию полноценных колоний) необходимо несколько
взаимодействий − теория многих ударов (n ударов). Близкая к ней
по форме теория многих мишеней рассматривает необходимость
одномоментным поражением нескольких мишеней в чувствительном
объеме клетки (n мишеней) для ее инактивации. В этом случае
кривые выживаемости не будут иметь строго экспоненциальный
характер, а образуют плечо.
Параметры, характеризующие радиочувствительность
клеток различных тканей
Ткань
Параметр D0,
Гр
Величина плеча
Dq, Гр
Стволовые кроветворные клетки костного мозга
(радиоустойчивая фракция)
0,9-1,0
1,5
радиочувствительная фракция
0,1
отсутствует
Крипты тонкой кишки
1,3
4,0 - 4,5
Стволовые клетки кожи
2,7
5,0
Эпителий почечных канальцев
1,5 - 1,6
3,9
Фолликулы щитовидной железы
2,0
2,3
Пролиферирующие эндотелиальные клетки
1,7 - 2,4
1,8 - 2,9
Клетки китайского хомячка, в различных фазах
клеточного цикла:
на стадии G1 и
в митозе
1,3
отсутствует
на стадии G1
1,6
n больше
1
на ранней стадии синтеза ДНК
1,9
n = 2,5
в поздней фазе синтеза ДНК
2,0
n = 10
Наиболее радиочувствительными среди всех видов клеток организма
являются лимфоциты, для которых величина параметра D0 составляет
0,5 Гр. Это характерно также для опухолевых клеток лимфоидного
происхождения.
Диапазон в области малых доз до 1 Гр
Диапазон в области малых доз до 1 Гр наиболее интересен с
позиции обеспечения радиационной безопасности, так как при
профессиональном облучении на Земле, при ликвидации радиационных
аварий наибольшее число лиц подвергается воздействию именно в
этих дозах.
Обнаружено, в частности, что кривая доза-эффект для ряда даже
радиоустойчивых линий клеток на начальном участке с ростом дозы
снижается относительно быстро, а затем следует плато, после чего
она принимает обычный вид. Повышенная радиочувствительность
клеток в диапазоне 0,1-0,5 Гр выявлена у 38 из 50 клеточных
линий, полученных из опухолей различных локализаций, нормальных
фибробластов и эпителиальных клеток человека. Такой же характер
инактивации мы наблюдали для стволовых кроветворных клеток при
экзоклонировании в селезенке.
Рассматриваемый диапазон доз характерен и для радиационного
воздействия космических излучений на космонавтов при
осуществлении длительных орбитальных полетов. При межпланетных
полетах, учитывая стохастический характер солнечных протонных
событий, возможны более высокие уровни воздействия радиации на
космонавтов.
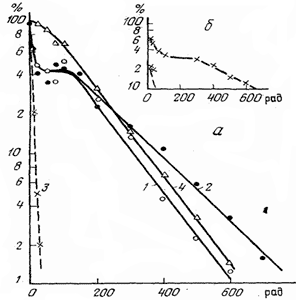
Рис.5. Выживаемость стволовых кроветворных клеток КОЕ (а) и
сперматогоний (б) у мышей:
1 – клонирование в селезенке; 2 – клонирование в костном мозге
бедренной кости; 3 – расчетная кривая для радиочувствительной
фракции КОЕ; 4 – клонирование в селезенке при дополнительном
введении Т-лимфоцитов
Поскольку ведущее значение в сохранении работоспособности и
жизнеспособности человека в ближайшем периоде в процессе
хронических и после кратковременных острых облучений принадлежит
кроветворной ткани, мы представим ниже основные зависимости от
дозы и мощности дозы сохраненной доли стволовых кроветворных
клеток, функциональных клеток в костном мозге и в периферической
крови.
На Рис. 5-а представлен характер относительного изменения числа
жизнеспособных стволовых кроветворных клеток (колоние-образующих
единиц (КОЕ)) у мышей от дозы острого гамма-облучения. Сходный
характер изменения числа жизнеспособных клеток имел место и при
облучении сперматогониев (Рис.5-б). Из данных рисунка видно, что при
малых значениях доз наблюдается резкое снижение числа стволовых
кроветворных клеток за счет радиочувствительной фракции (параметр D0 оказывается
равным 0,1 Гр). Затем форма кривой становится обычной, и последующее
снижение численности радиоустойчивой фракции клеток происходит по
многоударной модели с параметрами Doи Dq,
равными 1,0 Гр и 1,5 Гр, соответственно.
Снижение эффективности радиационного воздействия при
фракционированном облучении
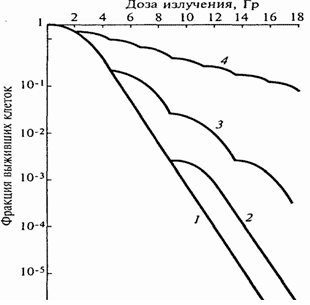
Рис.6. Кривые выживаемости клеток карциномы Эрлиха линии ELD,
растущих в культуре при фракционировании дозы радиационного
воздействия. Кривые 1 и 2 основаны на экспериментальных данных
однократного облучения и разделенного на две фракции; 3 и 4 –
расчетные кривые для разделения дозы 18 Гр на 4 и 8 равных фракций
на основе параметров кривой выживаемости для однократного облучения.
Большим числом радиобиологических исследований показано, что после
воздействия гамма или рентгеновского излучений и создании на несколько часов
условий для возможности восстановления поврежденной структуры ДНК отмечается
значительное увеличение доли выживших клеток. Это явление, обусловленное
созданием условий для активизации ферментативных систем репарации и
увеличением времени восстановления структур ДНК до начала синтеза и
вступления клеток в митоз, было названо восстановлением потенциально
летальных повреждений. В зависимости от условий скорость восстановления
определяется характеристическими временами от 1 до 10 часов.
На Рис.6 показано снижение эффективности радиационного
воздействия при фракционированном облучении. Увеличение доли выживших клеток
оказывается тем большим, чем больше число фракций и интервалы между ними.
Аналогичные процессы, связанные с быстрым восстановлением повреждений в
клетках и приводящие к резкому увеличению доли выживших клеток, имеют место
при разделении суммарной дозы на отдельные фракции с интервалами от
нескольких часов до нескольких суток. Этот тип восстановления был назван
восстановлением сублетальных повреждений, поскольку ослаблялись в этом
случае процессы закрепления поражения наиболее важных структур клеток,
уменьшался выход двойных разрывов ДНК и облегчались условия для
восстановления нормальных структур клеток до вступления части клеток в
наиболее радиочувствительные стадии.
Наиболее значимо увеличивается доля жизнеспособных клеток при
фракционировании радиационного воздействия для тканей с большими
возможностями восстановления сублетальных повреждений, кривая выживаемости
от дозы для которых характеризуется большими значениями плеча (параметра Dq).
Учет фактора восстановления радиационного поражения при облучении с
малыми значениями мощности дозы
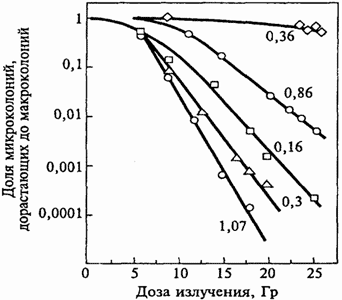
Рис.7. Выживаемость клеток китайского хомячка в зависимости от дозы
при снижении мощности дозы облучения с 1,07 Гр/мин до 0,36 сГр/мин.
Учет фактора восстановления радиационного
поражения на клеточном, тканевом и организменном уровнях важен не
только при фракционированном воздействии ионизирующих излучений, но
и при облучении с малыми значениями мощности дозы.
На Рис. 7 в качестве примера показано значительное снижение
поражения клеток китайского хомячка линии CHL –F при снижении
мощности дозы гамма-излучения от 1,07 до 0,16 Гр/мин и последующем
снижении до 0,36 сГр/мин. Здесь облучению подверглись клетки, уже
образовавшие микроколонии со средним числом клеток на колонию 3,5.
По оси ординат отложена доля микроколоний, дорастающая до
макроколоний. При совсем малых значениях мощности дозы снижение
эффективности облучения связано не только с восстановлением
сублетальных повреждений, но и с существенно большим временем
облучения. В этом случае воздействие осуществляется на все более
поздние поколения клеток.
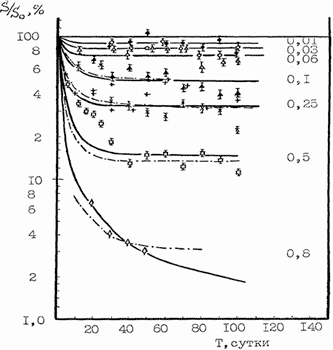
Рис.8. Относительное изменение числа стволовых кроветворных клеток
КОЕ во времени при протяженном гамма-облучении мышей с различной
мощностью дозы Гр/сут.
Такой же характер снижения эффективности радиационного воздействия
при уменьшении мощности дозы имеет место и для других
быстрообновляющихся тканей, в том числе и для кроветворной ткани.
Исследования выживаемости стволовых кроветворных клеток, образующих
колонии в селезенке КОЕ у мышей, показали, что при мощности дозы 50
сГр/час (около 1 сГр/мин) погибает в 3 раза меньше клеток, чем при
обычных мощностях острого облучения 50-100 сГр/мин.
Из данных Рис. 8 видно значительное снижение эффективности
радиационного воздействия и увеличение доли выживших стволовых
кроветворных клеток при снижении мощности дозы радиационного
воздействия.
Эффекты фракционирования дозы и мощности дозы необходимо учитывать
при оценке опасности для космонавтов в длительных полетах повторных
радиационных воздействий в результате реализации солнечных протонных
событий (СПС).
 На схеме отчетливо видно более глубокое радиационное повреждение
молекулы ДНК при действии ускоренных тяжелых ионов с высокими
линейными передачами энергии (ЛПЭ) по сравнению с воздействием
рентгеновского излучения.
На схеме отчетливо видно более глубокое радиационное повреждение
молекулы ДНК при действии ускоренных тяжелых ионов с высокими
линейными передачами энергии (ЛПЭ) по сравнению с воздействием
рентгеновского излучения.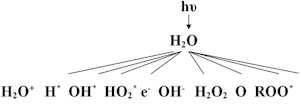
 Следует отметить, что если прямое повреждающее действие
осуществляется, можно считать, практически мгновенно, то
опосредованное действие носит существенно более длительный характер
с образованием порой токсичных белковых продуктов. При этом прямое
непосредственное действие ответственно лишь за 10-20 % лучевого
поражения.
Следует отметить, что если прямое повреждающее действие
осуществляется, можно считать, практически мгновенно, то
опосредованное действие носит существенно более длительный характер
с образованием порой токсичных белковых продуктов. При этом прямое
непосредственное действие ответственно лишь за 10-20 % лучевого
поражения.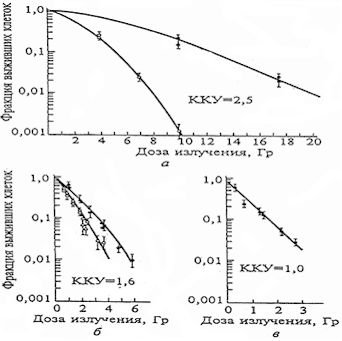
 Известно, что процесс ионизации структур клетки носит
стохастический характер. Для инактивации клетки при действии
плотноионизирующих излучений, как правило, бывает достаточно
реализации одного события попадания частицы в ядро клетки и
повреждения структуры ДНК. Кривые выживаемости клеток различных
тканей в этом случае описываются экспоненциальной зависимостью
от дозы:
Известно, что процесс ионизации структур клетки носит
стохастический характер. Для инактивации клетки при действии
плотноионизирующих излучений, как правило, бывает достаточно
реализации одного события попадания частицы в ядро клетки и
повреждения структуры ДНК. Кривые выживаемости клеток различных
тканей в этом случае описываются экспоненциальной зависимостью
от дозы: