Современное состояние проблемы цитогенетической индикации дозы
Опубликовано в журнале Медицинская радиология и радиационная безопасность
(http://www.medradiol.ru).
2016. № 5
Введение
M.L. Mendelsohn [1] в 1991 г. определил биологическую дозиметрию (биодозиметрию) как оценку физически, химически или биологически значимой дозы предшествующего воздействия, используя наблюдение за биологическим субъектом или осуществляя [физическими, химическими или биологическими способами] измерение параметров материала, полученного из этого субъекта. Правда, с нашей точки зрения, выражение «биологическая дозиметрия», вообще говоря, не является корректным, так как с физической точки зрения доза определяется как энергия, поглощённая единицей массы вещества. При облучении же любая измеряемая в биологическом объекте реакция зависит не только от этого параметра, но и от характера излучения, мощности дозы и распределения её во времени, состояния самого биологического субстрата. Поэтому более правильно говорить о биологической индикации дозы в той мере, в какой её величина отражается в соответствующих исследованиях. Поэтому термин «биодозиметрия», с нашей точки зрения, следует использовать исключительно в целях фразеологического разнообразия.
При этом собственно физическая дозиметрия, безусловно, обладает более высокой точностью и чувствительностью. Однако эти ее преимущества могут быть воплощены в жизнь только тогда, когда сама физическая дозиметрия является доступной. На практике же в различных аварийных ситуациях индивидуальные дозиметры чаще всего отсутствуют или не вполне адекватны. Это характерно при вовлечении в аварии как профессиональных работников, так и, особенно, тех людей, кто по роду своей деятельности и образу жизни не должны подвергаться сверхфоновому радиационному воздействию (исключая медицинские процедуры).
Одновременно, как правило, не достает и достоверных сведений
о длительности облучения, мощности дозы, расстоянии между пострадавшим и источником излучения, положении тела в период радиационного воздействия, которые позволяли бы производить соответствующие дозиметрические расчеты. Например, по данным Национального совета по радиационной защите (National Radiological Protection Board)
Великобритании из 904 ситуаций с подозрением на переоблучение, имевших место с 1968 г. по 1996 г., в 280 случаях какие-либо сведения по физической дозиметрии вообще отсутствовали [2].
В этих условиях единственными источниками необходимых данных
становятся методы биологической индикации дозы. Отметим также мнение В.Г.Владимирова [3], который считает, что индивидуальные особенности функционального состояния или конституции конкретного организма, включая его радиочувствительность, могут существенно сказаться на степени тяжести и последствиях поражения. следовательно, предположение о том, что биологические индикаторы позволяют определенным образом учитывать эти обстоятельства, отражая одновременно и величину реальной («физической») поглощенной дозы, является довольно обоснованным.
Цитогенетическая оценка дозы после острого облучения
M.A. Bender и P.C. Gooch в 1962 г. предложили использовать подсчёт аберраций
хромосом, индуцированных в лимфоцитах периферической крови после случайного
облучения людей, для дозовых оценок с помощью построенных на основании опытов in vitro кривых «доза−эффект» [4, 5]. В настоящее время цитогенетический анализ культур лимфоцитов периферической крови является одним из основных способов биологической индикации дозы, что отражено в методических рекомендациях МАГАТЭ
(2011 г.) [6]. Так как подавляющая часть лимфоцитов в крови находится в фазе G0 (покоя) клеточного цикла, то при радиационном воздействии в них происходит индукция аберраций хромосомного типа.
Несмотря на определённые трудности, связанные с характером и режимом воздействия, анализ аберраций хромосом достаточно хорошо зарекомендовал себя в случаях аварийного относительно равномерного облучения в дозах, вызывающих развитие острой лучевой болезни (ОЛБ)
(1 Гр и более). Как известно, в системе культур, стимулированных in vitro к делению лимфоцитов периферической крови, основным цитогенетическим индикатором радиационного воздействия считаются дицентрики. анализ этого вида аберраций хромосом даже называют «золотым стандартом» для биологической дозиметрии [7–9], что обусловлено их характерным внешним видом и низкой спонтанной частотой.
Хотя наблюдается значительная вариабельность данных в различных исследованиях, но в среднем частота дицентриков составляет 1 на 1000 проанализированных клеток. Эта величина принята как норма в рекомендациях МАГАТЭ [6], хотя, по данным отдельных авторов, она может колебаться в достаточно широких пределах. Так, в статье [10], посвящённой сравнению фоновых
уровней повреждений хромосом, обнаруженных разными отечественными цитогенетическими лабораториями, средние частоты дицентриков (на 100 клеток) в возрастной группе 18 лет и старше варьировали от 0,005 до 0,143.
В настоящее время для стандартизации данных, получаемых в радиационной цитогенетике, предприняты серьёзные усилия для создания баз данных с электронными изображениями метафаз с дицентриками,
которые были предварительно изучены международными экспертами. Например, в хранилище международной сети лабораторий BioDoseNet, занимающихся цитогенетической индикацией дозы, сейчас содержится примерно 25 тыс. таких изображений,
которые могут быть использованы как в учебных, так и исследовательских целях [11].
Также большое значение имеет хорошо выраженная дозовая зависимость для радиационно-индуцированной частоты дицентриков. При этом для оценки дозы редкоионизирующих и плотноионизирующих излучений используются кривые «доза-эффект» линейноквадратичного и линейного вида соответственно для средней частоты этих аберраций, полученные по результатам облучения крови здоровых доноров
in vitro, так как радиочувствительность хромосом лимфоцитов одинакова при облучении в организме и вне него. При этом каждой лаборатории рекомендуется иметь собственные кривые «доза-эффект» в основном в связи с особенностями идентификации аберраций хромосом и выбора метафаз для анализа.
Альтернативным подходом может служить оценка не самой дозы, а величины гамма-эквивалентного биологического повреждения по дозовым кривым, полученным при гамма-облучении
in vitro. В этом есть смысл, например, при предсказании конечной тяжести радиационного поражения по гематологическим данным. Так, в нашей лаборатории наблюдалась сильная корреля‑ ция между цитогенетическим оценками дозы и её оценками по «стандартным» кривым пострадиационной динамики числа нейтрофилов в периферической крови, которые были получены для редкоионизирующих излучений, у лиц, пострадавших при аварии на Чернобыльской
АЭС и имевших клинические проявления ОЛБ [12].
Отметим, что дицентрики относятся к, так называемым, нестабильным аберрациям хромосом, т.е. имеют тенденцию к элиминации с течением времени после облучения в силу того, что могут представлять механическое препятствие для протекания митоза. Поэтому для биологической индикации дозы необходимо производить цитогенетический анализ только или главным образом в клетках первого митоза в культуре. В своё время была предложена методика дифференциального окрашивания сестринских хроматид. другие названия: арлекиновая окраска или окраска флуоресцент + Гимза (кратко FPG‑окраска). Она позволяет различать метафазы первого (однородная окраска хромосом), второго и последующих митозов в культуре, вообще говоря, любых клеток. Поэтому для исследования частот аберраций только в лимфоцитах первого митоза было предложено использовать данный метод. альтернативный подход заключается в сокращении сроков культивирования насколько возможно с тем, чтобы в культуре присутствовали преимущественно клетки первого митоза, что, конечно, надо исходно подтвердить в предварительных исследованиях.
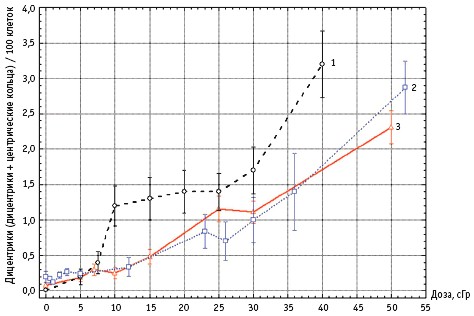
Порогом чувствительности метода для редкоионизирующих излучений при использовании классической окраски хромосом считается доза 0,1 Гр острого воздействия [6]. При этом одной из проблем биологической индикации дозы остаётся её оценка в «субклиническом» диапазоне, при котором не происходит развития ОЛБ (т.е. меньше 1 Гр). до сих пор существуют разночтения в ходе дозовой зависимости при таком уровне поражения, так как ряд исследователей обнаруживали плато в выходе дицентриков
при определённых дозах (100–300 мЗв) [13], тогда как другие этого не наблюдали
[14, 15]. На рис. 1 эти данные представлены графически. авторы [14],
категорически отрицающие наличие закономерных отклонений от монотонно
возрастающей дозовой функции, связывают их феноменологическое проявление со
слабой статистикой из-за недостаточного числа проанализированных метафаз при общем низком уровне индуцированных аберраций хромосом. При этом заметим, что в работе [14], в которой были использованы очень низкие дозы, частоты аберраций хромосом при дозах меньше 10 мЗв были вообще ниже фоновых значений. В плане цитогенетической оценки доз менее100 мЗв интересна работа [7]. В ней было обследовано 10 пациентов с различными заболеваниями до и после проведения компьютерной томографии (КТ). Полученные физические дозы варьировали от 5,78 до 60,27 мЗв. При цитогенетическом анализе (по 2000 метафазы) были использованы два подхода: классический (Гимза) метод и метод флуоресцентной гибридизации
in situ (FISH) с использованием ДНК-зондов к центромерам хромосом. В обоих вариантах наблюдалось значимое групповое превышение частот дицентриков после КТ над их допроцедурным уровнем. Однако, во-первых, не всегда это превышение было существенным у отдельных пациентов и, во-вторых, отсутствовала дозовая зависимость.
Авторы рассуждают о возможности увеличения чувствительности обоих методов путём увеличения числа проанализированных клеток (до 10 тыс. на человека). Однако мы испытываем большие сомнения по этому вопросу и склонны считать, что принятая МАГАТЭ нижняя граница чувствительности цитогенетического метода (100 мЗв) объективно более соответствует действительности при индивидуальных оценках дозы.
Рис. 1. результаты определения различными авторами частот дицентриков (кривая 1 – [13], кривая 2 – [14]) или дицентриков + центрических колец (кривая 3 – [15]) в культурах лимфоцитов периферической крови здоровых доноров после гамма-облучения
in vitro в дозах до 1 Гр
В настоящее время в связи с развитием различных вариантов FISH‑метода действительно появилась возможность более точного учёта дицентриков при одновременном локальном FISH‑окрашивании в разные цвета цетромерных и теломерных участков хромосом, например, в зелёный и красный цвета соответственно. В этом случае моноцентрическая метафазная хромосома имеет один зелёный центромерный сигнал и 4 красных теломерных сигнала; дицентрик характеризуется 2 зелёными сигналами и 4 красными сигналами; у центрического кольца выявляется один зелёный сигнал без красных, а у ацентрического кольца нет никаких сигналов; парные фрагменты имеют 2 или 4 красных сигнала. В работе M’kacher et al.
[8] такой подход привёл к увеличению частоты регистрируемых дицентриков в целом до +30 %. Правда, парадоксальным образом, такое увеличение было выражено в контроле и при дозах 0,5 Гр и больше, а при дозах 0,1 и 0,2 Гр его не наблюдалось. Поэтому необходимы дальнейшие исследования.
Рис. 2. автоматизированная система «Метафер4» для автоматического поиска метафаз, кариотипирования и проведения FISH‑исследований (фирма «Метасистемс», цитогенетическая оценка неравномерности радиационного воздействия
Следует отметить, что в настоящее время имеются возможности значительно ускорить и облегчить работу цитогенетиков
в связи с практическим применением устройств по автоматизированному цитогенетическому анализу. с этой целью была разработана, например, система «Метафер» фирмы «Метасистемс» (Германия).
Её первоначальное компьютерное обеспечение позволяло только производить автоматическое сканирование предметных стёкол с целью поиска метафаз [15]. В настоящее время имеются возможности полуавтоматического анализа дицентриков, правда, требующего внешнего контроля [16]. При этом конечная задача состоит в достижении полностью автоматического цитогенетического определения числа дицентриков
и построении на его основе соответствующих калибровочных кривых
[17]. Важную роль играет соответствующий вариант компьютерного
обеспечения системы «Метафер» при проведении различных FISH‑исследований. В ряде учреждений
России этот компьютерно‑микроскопический комплекс различных поколений уже применяется. На рис. 2 представлена фотография научного сотрудника ФМБЦ им.
А.И. Бурназяна М.Г. Козловой, занимающейся цитогенетическими
исследованиями с привлечением описываемой автоматизированной системы.
Цитогенетическая оценка неравномерности радиационного
воздействия
При облучении в дозах, вызывающих ОЛБ, существенной проблемой для прогнозирования её тяжести является наличие или отсутствие существенной неоднородности облучения. Как известно, для качественного её выявления в радиационной цитогенетике используется изучение распределения клеток по числу содержащихся в них дицентриков в культуре лимфоцитов периферической крови.
Его соответствие теоретическому распределению Пуассона свидетельствует
об относительной равномерности радиационного поражения, отклонение в сторону большей дисперсии – о неравномерности распределения дозы по массе тела.
В методических рекомендациях МАГАТЭ [6] предлагаются два подхода для оценки доли пораженного участка и поглощённой в нём дозы при парциальном радиационном воздействии, обозначаемые как «загрязнённый» метод Dolphin и Qdr‑метод. Оба способа требуют компьютерного сопровождения и ограничены вариантом, когда в одной определённой дозе облучена одна часть тела, а другая остаётся непоражённой. ранее нами был предложен более общий подход оценки распределения доз по массе тела при разных вариантах неравномерного облучения, основанный на специальной компьютерной программе ныне покойного доктора биол. наук
И.В. Филюшкина и представлении, что в смеси клеток, облучённых в разных дозах, распределение дицентриков по клеткам является суперпозицией парциальных пуассоновских распределений [19]. В статье [20] была продемонстрирована работа этой программы на примере
восстановления с её помощью дозовых распределений после составления
искусственных («на бумаге») смесей лимфоцитов, облучённых в разных
дозах. Использовались различные варианты, и в целом наблюдалось достаточно близкое совпадение заданных и восстановленных распределений доз.
Однако кроме использования самого компьютерного метода, необходимо учитывать дозо‑зависимые процессы интерфазной
гибели лимфоцитов и задержки их пролиферации. с этой целью нами были проведены эксперименты со смешанными культурами лимфоцитов из подвергнутой и не подвергнутой
in vitro гамма‑облучению (50 на 50 %) крови здоровых доноров [21]. После применения компьютерной программы к наблюдаемым распределениям дицентриков в чисто облучённых и в смешанных культурах лимфоцитов, как и ожидалось, культуры облученных лимфоцитов демонстрировали 100 %‑ую фракцию облученных клеток, а оценки доз на облученную фракцию были близки к дозам, определенным по частоте дицентриков. В смешанных же культурах восстановленное
значение фракции облученных клеток зависело от дозы. При дозах 1 и 2
Гр оно оказалось несколько выше заданного значения 50 %, и близким к нему при дозе 3 Гр. с ростом дозы наблюдалась дальнейшая тенденция к снижению доли облученных клеток. При наивысшей из использованных доз (8 Гр) восстановленное значение фракции облученных клеток равнялось примерно 30 %, а не заданным – 50 %. В целом значения восстановленной дозы на облученную фракцию в смешанных культурах были всегда несколько ниже, чем в культурах облученных лимфоцитов, что особенно видно при дозе 8 Гр.
Нами были предложены соответствующие уравнения для коррекции как объема облученной фракции, так и величины оценки дозы в случаях неравномерного облучения.
Также в Клиническом отделе бывшего института биофизики Минздрава
СССР (ныне – ФМБЦ им. А.И. Бурназяна ФМБА России) были разработаны
методики оценки поглощённых доз на любые участки костного мозга, доступные для пункции, по числу аберрантных клеток в «прямых» препаратах и по частотам аберраций хромосом в культурах лимфоцитов костного мозга [19]. Ниже приведено следующее уравнение для индикации дозы согласно первому методу:
D = (2 arcsin √P/100 – 1,445 – 0,177×T) / 0,286,
где D – поглощённая доза, Гр; Р – процент аберрантных клеток; Т – время,
прошедшее после облучения, ч. Последний показатель необходим для учёта
элиминации аберрантных клеток в связи с высокой пролиферативной активностью большинства клеточных элементов гемопоэтической ткани. На рис. 3 представлено графическое решение данного уравнения в виде номограммы. Отметим, что, так как в костном мозге клетки находятся в различных фазах клеточного цикла, то облучение индуцирует в них аберрации как хромосомного, так и хроматидного типов.
Рис. 3. Номограмма для оценки локальной дозы (D, Гр) в пунктатах костного мозга в зависимости от процента (%) аберрантных клеток в «прямых» препаратах хромосом и времени (Т, ч), прошедшего после облучения (f = 1,445 + 0,286×D – 0,0177×T)
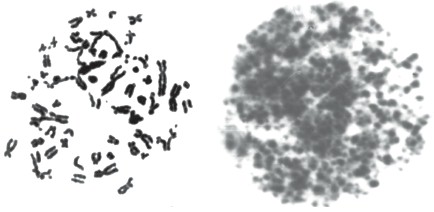
Однако возможность дозовых оценок по D данной схеме не превышает 4 сут после воздействия, так как при больших сроках обнаруживаемая частота аберрантных клеток может не отличаться от спонтанных значений. Также имеется дозовое ограничение в связи с тем, что частота аберрантных клеток при анализе через 24 ч после облучения достигает 100 % уже при дозе 5 Гр, а подсчёт общего числа аберраций становится невозможным в отдельных метафазах
уже при дозе 4 Гр, и с возрастанием дозы количество таких клеток увеличивается. Поэтому при больших дозах возможна только экспертная оценка. На рис. 4 для примера представлены такие метафазные клетки из пунктатов
облучённого костного мозга. если в метафазе слева ещё различимы хромосомные структуры, то в метафазе справа наблюдается полная фрагментация хромосом.
Для лимфоцитов костного мозга ранее в нашей лаборатории было показано, что их радиочувствительность не отличается от радиочувствительности лимфоцитов периферической крови [22]. использование для индикации локальных
доз цитогенетического анализа культур лимфоцитов костного мозга из его различных участков ограничивается разведением пунктатов периферической кровью. соответственно, при исследовании наиболее и наименее поражённых участков кроветворной ткани будут наблюдаться занижение и завышение дозы соответственно.
Следует признать, что все представленные выше методы цитогенетической оценки неравномерности облучения имеют свои преимущества и недостатки, и поэтому они должны использоваться комплексно.
Рис. 4. Клетки в «прямых» препаратах пунктатов костного мозга у случайно облучённых пациентов со средней дозой на всё тело 5,5 Гр (фото слева) и примерно 9 Гр (фото справа)
Ретроспективная цитогенетическая оценка дозы
Другой задачей всегда стоявшей и стоящей перед любой
биологической индикацией дозы является её ретроспективная оценка. Примерно до последнего двадцатилетия ХХ века для решения этой проблемы с помощью различных подходов пытались использовать классическую окраску хромосом, несмотря на то, что она позволяет в основном анализировать нестабильный тип аберраций хромосом (к ним относятся и дицентрики), которые имеют тенденцию элиминировать с течением времени. G‑бэндинг,
широко применяемый в онкогематологии для полногеномного кариотипирования и выявления клоновых стабильных перестроек хромосом, оказался слишком трудоёмким для достижения данной поставленной цели.
Только после внедрения FISH‑окрашивания целых хромосом появился относительно лёгкий для анализа способ учёта реципрокных транслокаций,
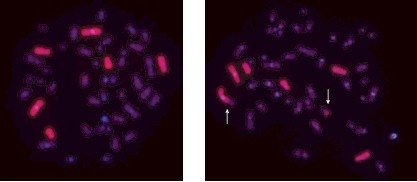
не представляющих механического препятствия для протекания митоза и теоретически имеющих тенденцию сохраняться с течением времени после облучения. На рис. 5 в качестве примера представлены фотографии двух метафаз (нормальной и аберрантной) из культуры лимфоцитов периферической крови пациента (№ 1094, первоначально оцененная доза по частоте дицентриков 6,6 Гр), пострадавшего при аварии на Чернобыльской
АЭС и повторно поступившего в клинику ФМБЦ им. А.И. Бурназяна для обследования через 25 лет
Рис. 5. FISH‑окрашенные (1, 4 и 12 пары хромосом) метафазы из культуры лимфоцитов периферической крови пациента (№ 1094, первоначальная оценка дозы по частоте дицентриков – 6,6 Гр), пострадавшего при аварии на Чернобыльской
АЭС, через 25 лет после облучения: слева – нормальная метафаза; справа – метафаза с транслокацией, в которой участвует 1 хромосома (транслоцированные участки показаны стрелками) после облучения, при FISH‑окрашивании препарата его хромосом с использованием
ДНК‑зондов к 1, 4 и 12 парам хромосом.
Основной принцип, который изначально закладывался в FISH‑ретроспективную
оценку дозы, заключался в использовании калибровочной дозовой зависимости для реципрокных транслокаций после острого облучения крови здоровых доноров
in vitro аналогично тому, как это делается для оценки дозы по частоте дицентриков. Однако по результатам цитогенетических исследований в отдалённые сроки после аварий на Чернобыльской
АЭС (1986 г.) [23] и в г. Гойянии (Бразилия, 1987 г.) [24] оказалось, что такой подход напрямую работает только до доз 1–2 Гр.
Действительно, при больших дозах частота регистрируемых FISH‑транслокаций становилась меньше первоначальной частоты дицентриков (в предположении их равновероятной индукции), что, по‑видимому, было обусловлено совместной элиминацией стабильных и нестабильных аберраций при их нахождении в одних и тех же клетках.
Такая зависимость породила представление о необходимости учёта FISH‑транслокаций
не во всех клетках, а только в стабильных, т.е. содержащих исключительно аберрации стабильного типа. На практике для большинства случаев ретроспективной оценки дозы в диапазоне до 1 Гр, когда в аберрантных
клетках содержится преимущественно по 1 перестройке хромосом, вполне достаточно
использования исходного подхода. Однако существенным является то, что в целом с
возрастом в контрольных популяциях увеличивается и средняя частота FISH‑регистрируемых транслокаций [25–27]. При этом возрастает и размах колебаний индивидуальных значений. Таким образом, порождается неопределённость при оценке дозы с помощью FISH‑методики, так как неизвестной оказывается истинная величина присущей данному человеку фоновой частоты транслокаций. Кроме того, в работе [26] продемонстрировано явное различие уровней транслокаций, регистрируемых в лабораториях из четырёх различных регионов нашей планеты. Поэтому встаёт вопрос о продолжении цитогенетических исследований в контрольных популяциях.
Для ретроспективной оценки дозы рутинным стало применение одноцветной FISH‑окраски 3 пар крупных хромосом (по выбору из групп
А, В и С) с последующим пересчётом на весь геном по специальной формуле [6] обнаруженной частоты обменов участками между окрашенными и неокрашенными хромосомами с учётом относительного содержания
ДНК в выбранных хромосомах. Причём рекомендуется использовать только полные реципрокные транслокации,
так как для них показана большая устойчивость во времени после облучения в отличие от неполных транслокаций. Последние могут быть результатом потери (делеции) терминального ацентрического фрагмента, т.е. клетка начинает относиться к потенциально нестабильному типу вследствие возникновения несбалансированного набора хромосом. Правда, часть перестроек, визуально идентифицируемых как неполная транслокация, являются результатом обмена с участием малоразмерных участков хромосом, лежащих за границами разрешения FISH‑метода. Поэтому, с нашей точки зрения, неучёт «псевдонеполных» транслокаций может оказаться не такой уж и правильной политикой.
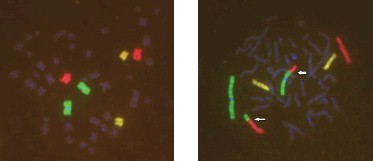
Для наблюдения за транслокациями между окрашенными хромосомами предлагается использовать так называемую трёхцветную FISH‑окраску [28, 29]. для пересчёта на весь геном при этом используется своя специальная формула [6]. Предполагается, что данный вариант FISH‑окрашивания может увеличить чувствительность метода при ретроспективной оценке дозы. На рис. 6, аналогично рис. 5, показаны нормальная и аберрантная метафазы у другого пациента (№ 1047, первоначально оцененная доза по частоте дицентриков 3,2 Гр), пострадавшего при аварии на Чернобыльской
АЭС, через 29 лет после облучения. Однако в данном случае был использован трёхцветный FISH‑метод: 1 пара хромосом имеет зелёный цвет, 4 пара хромосом – красный цвет и 12 пара хромосом – жёлтый цвет. Таким образом, выявлена транслокация между 1 и 4 парами хромосом, которая при одноцветном FISH‑окрашивании оказалась бы нераспознанной.
Рис. 6. Трёхцветный FISH‑метод (1 пара хромосом – зелёный цвет, 4 пара хромосом – красный цвет, 12 пара хромосом – жёлтый цвет): метафазы из культуры лимфоцитов периферической крови пациента (№ 1047, первоначально оцененная доза по частоте дицентриков – 3,2 Гр), пострадавшего при аварии на Чернобыльской
АЭС, через 29 лет после облучения: слева – нормальная метафаза; справа – метафаза с транслокацией между1 и 4 хромосомами
Однако наиболее полный учёт транслокаций, о чём говорится в рекомендациях
МАГАТЭ [6], возможен при использовании методик, обеспечивающих разноцветное окрашивание (после компьютерной обработки изображений) всех пар хромосом: mFISH и SKY (спектральное кариотипирование). Однако полноценное использование этого подхода требует исследования фоновых значений регистрируемых при этом частот аберраций хромосом и построения соответствующих дозовых кривых.
На самом деле основной причиной разработки выше упомянутых многоцветных вариантов FISH в большей степени являлось стремление выявить сложные межхромосомные аберрации. Под этим термином понимают ситуацию, когда количество разрывов хромосом, по крайней мере, на один больше, чем общее число хромосом, участвующих в данной общей перестройке. Такие аберрации могут быть обнаружены даже при действии редкоионизирующих излучений, что не регистрируется при классическом окрашивании. Но наиболее характерны они для плотноионизирующих излучений (нейтроны, альфа‑частицы, тяжёлые ионы). Лимфоциты с такими аберрациями были обнаружены, например, у работников плутониевого производства, наблюдавшихся в клинике ФМБЦ им.А.И.Бурназяна [30]. Однако по большей части эти клетки
относились к нестабильному, т.е. имеющему затруднения к передаче в дочерние клетки в процессе пролиферации, типу, хотя и могли служить индикатором хронического альфа‑облучения. Более последовательное изучение проблемы было выполнено на пробах крови работников ПО «Маяк» [31, 32]. В этом исследовании были проанализированы стабильные клетки со сложными и множественными (т.е. несколько простых обменов в одной метафазе) транслокациями. их повышенная частота наблюдалась у работников с высоким содержанием плутония в организме, но не у тех лиц, которые подверглись воздействию только внешнего γ‑облучения.
В научной литературе также содержится утверждение, что для действия плотноионизирующих излучений более надёжным показателем, чем межхромосомные, являются внутрихромосомные
перестройки. для обнаружения последних может быть использована ещё одна FISH‑методика, обозначаемая как mBAND, при применении которой в выбранной паре хромосом их отдельные участки, расположенные вдоль длины хромосом, приобретают разную окраску. На основе этого подхода было, например, показано, что альфа‑излучение создаёт гораздо больше внутрихромосомных обменов, чем редкоионизирующее излучение, что предположительно создаёт возможность для их различения при биологической индикации дозы [33]. с помощью этой технологии было проведено цитогенетическое обследование работников ПО «Маяк» и установлена линейная корреляция частоты внутрихромосомных
обменных аберраций с поглощённой дозой внутреннего альфа‑облучения и содержанием
239Pu в организме (коэффициенты корреляции 0,69 и 0,76 соответственно) [34], хотя необходимо обратить внимание на имевшийся широкий индивидуальный разброс. Правда, в более поздней работе те же авторы (Н.В.
Сотник и Т.В.Азизова) [35] сравнили использование mBAND и mFISH методов также при обследовании работников ПО «Маяк» и пришли к выводу, что они в принципе равноценны при биологической индикации действия инкорпорированных альфа‑излучателей. если же имеет место сочетанное воздействие, метод mFISH
более информативен, так как позволяет различать маркеры внешнего гамма‑излучения (простые транслокации)
и внутреннего альфа‑излучения (сложные перестройки хромосом).
Метод преждевременной конденсации хромосом
Кроме описанных выше цитогенетических методик, для биологической индикации дозы был разработан метод преждевременной конденсации хромосом (premature chromosome condensation – PCC). Первоначальный вариант (который используется и сейчас) состоял в слиянии (например, под действием инактивированного вируса сендай или полиэтиленгликоля) интерфазных лимфоцитов периферической крови человека с делящимися в культуре клетками млекопитающих или клетками HeLa [36–38]. В результате происходила определённая конденсация интерфазных человеческих хромосом. Этот метод технически достаточно труден, а индекс конденсации низок.
Позднее был предложен химический способ рсс, индуцированной ингибиторами серин/треонин протеиновых фосфатаз типов 1 и 2А (окадаиновая кислота и калликулин а) [6]. В большинстве вариантов требуется стимуляция лимфоцитов к делению ФГА, но также сообщалось и о методике без такого воздействия [39]. В последнем случае избегается процедура длительного культивирования и, соответственно, снижается влияние интерфазной гибели и митотической задержки.
Сам РСС индуцируется практически в любой фазе клеточного цикла, хотя сейчас чаще применяют анализ в G2/M.
Этот подход наиболее полезен при высоких дозах редкоионизирующих излучений, а также при действии плотноионизируюших излучений и имеет несколько
вариаций, при которых могут учитывать фрагменты или кольца (окраска по Гимза), дицентрики (С‑бэндинг), транслокации (FISH‑окрашивание) [40–49].
Дозовые зависимости при этом чаще имеют линейный характер, однако могут выходить на плато при очень больших дозах. Также идёт разработка различных способов автоматизации подсчёта аберраций хромосом при использовании
РСС‑метода [50, 51].
Заключение
В данный небольшой обзор не вошли такие группы методов как анализ микроядер и тесты
ДНК‑комет. Хотя эти подходы также относятся к цитогенетическим методикам, но, во‑первых, мы ими практически не занимаемся и, во‑вторых, они сами по себе заслуживают отдельного обсуждения.
Относительно же рассмотренных вопросов, мы хотим сделать
следующее заключение. имеющиеся в настоящее время методики биологической индикации дозы с использованием аберраций хромосом очень разнообразны. Классический метод остаётся основным при оценке дозы в ближайшие сроки после облучения. спорными остаются вопросы о ходе дозовой зависимости при дозах ниже 1 Гр и о нижней границе чувствительности цитогенетических методик (как классической, так и FISH). В рекомендациях
МАГАТЭ для стандартного метода она считается равной 100 мГр, хотя имеются сообщения о её снижении до 50 мГр и даже до 20 мГр. имеется неопределённость в величине фоновой частоты FISH‑регистрируемых транслокаций у отдельных людей, связанная с её зависимостью от возраста и использованием при расчётах доз средних фоновых частот, характерных для того или иного возраста.
Использование многоцветных вариантов FISH, по‑видимому, будет способствовать повышению чувствительности и точности метода при ретроспективной оценке дозы, хотя для его полноценного использования потребуется изучение фоновых частот в различных популяциях.
Усовершенствование РСС‑методики, с нашей точки зрения, будет
способствовать расширению диапазона, в рамках которого будет возможна надёжная биологическая индикация дозы. Как уже известные, так и новые подходы будут находить своё место в различных ситуациях. В целом в настоящее время характерным является соединение различных цитогенетических методик в ту или иную одну технологию для решения соответствующих поставленных задач.
СПИСОК ЛИТЕРАТУРЫ
Mendelsohn M.L. Introduction to biological dosimetry // In: New Horizons in Biological Dosimetry. Proc. Internat. Symp. on Trends in Biological Dosimetry, held in Lerici, October 23–27, 1990. Eds. B.L. Gledhill, F. Mauro. – New York: Wiley‑Liss, Inc. 1991. P. 1–10.
Lloyd D., Edwards A., Moquet J. et al. Doses in radiation accidents investigated by Chromosome aberration analysis. XXI. Review of cases investigated, 1994–1996. – Chilton: National Radiological Protection Board. 1996. 24 pp.
Владимиров В.Г. Предисловие // Радиобиологические подходы к
диагностике лучевых поражений (Сборник научных трудов). – Л. 1987. C. 3–5.
Bender M.A., Gooch P.C. Persistent chromosome aberrations in irradiated human subjects // Radiat. Res. 1962. Vol. 16. № 1. P. 44–53.
Bender M.A., Gooch P.C. Types and rates of X‑ray‑induced chromosome aberration yields in human blood irradiated in vitro // Proc. Nat. Acad. Sci. U.S.A. 1962. Vol. 48. № 4. P. 522–532.
Cytogenetic dosimetry: applications in preparedness for and response to radiation emergencies – Vienna: IAEA. 2011. 245 pp.
Abe Yu., Miura T., Yoshida M.A. et al. Increase in dicentric chromosome formation after a single CT scan in adults // Scientifc Reports. 2015. № 5: 13882. 9 p. DOI: 10.1038/ srep13882.
M’kacher R., Maalouf E.E.L., Ricoul M. et al. New tool for biological dosimetry: Reevaluation and automation of the gold standard method following telomere and centromere staining // Mutat. Res. 2014. Vol. 770. № 1. P. 45–53.
Sullivan J.M., Prasanna P.G.S., Grace M.B. et al. Assessment of biodosimetry methods for a mass‑casualty radiological incident: medical response and management considerations // Health Phys. 2013. Vol. 105. № 6. P. 540–554.
Севанькаев А.В., Хвостунов И.К., Снигирёва Г.П. и соавт. Сравнительный анализ результатов цитогенетических обследований контрольных групп лиц в различных отечественных лабораториях // Радиац. биол. Радиоэкология. 2013. Т. 53. № 1. С. 5–24.
Romm H., Beinke C., Garcia O. et al. A new cytogenetic biodosimetry image repository for the dicentric assay // Radiat. Protect. Dosim.. 2016. 9 p. Advance Access published July 13, 2016. doi:10.1093/rpd/ncw158.
1Чирков А.А., Чистопольский А.С., Нугис В.Ю. О влиянии
мощности дозы на прогноз динамики количества нейтрофилов в крови по результатам цитогенетического анализа лимфоцитов у лиц, пострадавших при аварии на Чернобыльской АЭС // Мед. радиология. 1991. Т. 36. № 1. С. 36–38.
Luchnic N.V., Sevankaev A.V. Radiation‑induced chromosomal aberrations in human lymphocytes. I. Dependence on the dose of gamma‑rays and an anomaly at low doses // Mutat. Res. 1976. Vol. 36. № 3. P. 363–377.
Lloyd D.C., Edwards A.A., Léonard A. et al. Frequencies of chromosomal aberrations induced in human blood lymphocytes by low doses of X‑rays // Int. J. Radiat. Biol. 1988. Vol. 53. № 1. P. 49–55.
Huber R., Kulka U., LÖrch T. et al. Automated metaphase fnding: an assessment of the efciency of the METAFER2 system in a routine mutagenicity assay // Mutat. Res. 1995. Vol. 334. Issue 1. P. 97–102.
Sun M., Moquet J.E., Barnard S. et al. Doses in radiation accidents investigated by chromosomal aberration analysis XXV. Review of cases investigated, 2006–2015. PHE‑CRCE‑025. – Chilton, Didcot: Centre for Radiation, Chemical and Environmental Hazards, Public Health England. 2016. 26 pp.
Rogan P.K., Li Y., Wilkins R. et al. Radiation dose estimation by automated cytogenetic biodosimetry // Radiat. Prot. Dosim. 2016. 11 pp. Advance Access published July 13, 2016. doi:10.1093/rpd/ncw161.
Снигирева Г.П., Хаймович Т.И., Шевченко В.А. Использование цитогенетических методов для биологической дозиметрии. Методические рекомендации, РФЯЦ‑ВНИИЭФ, РНЦ рентгенорадиологии МЗ РФ, Институт общей генетики РАН. – Саров: РФЯЦ‑ВНИИЭФ. 2003. 55 с.
Пяткин Е.К., Баранов А.Е., Филюшкин И.В. и др. Оценка дозы и равномерности облучения при острых радиационных поражениях человека с помощью анализа аберраций хромосом. Методические рекомендации. – М.: Минздрав СССР. 1988. 25 с.
Пяткин Е.К., Филюшкин И.В., Нугис В.Ю. Оценка равномерности
облучения по результатам цитогенетического исследования лимфоцитов периферической крови человека // Тер. архив. 1986. Т. 58. № 9. С. 30–33.
Филюшкин И.В., Нугис В.Ю., Чистопольский А.С. Сравнительный цитогенетический анализ культур облучённых лимфоцитов и смешанных культур облучённых и необлучённых клеток // Мед. радиол. и радиац. безопасность. 1999. Т. 44. № 3. С. 19–26.
Пяткин Е.К., Покровская В.Н., Триска В.В. Частота хромосомных аберраций в культурах лимфоцитов костного мозга и периферической крови человека после γ‑облучения in vitro // Мед. радиология. 1980. Т. 25. № 2. С. 44–48.
Sevan’kaev A.V., Khvostunov I.K., Mikhailova G.F. et al. Novel data set for retrospective biodosimetry using both conventional and FISH chromosome analysis afer high acci‑ dental overexposure // Appl. Radiat. Isotopes. 2000. Vol. 52. № 5. P. 1149–1152.
Natarajan A.T., Santos S.J., Darroudi F. et al. 137Cesium‑induced chromosome aberrations analysed by fuorescence in situ hybridization: Eight years follow up of the Goiậnia radia‑ tion accident victims // Mutat. Res. 1998. Vol. 400, № 1. P. 299–312.
Воробцова И.Е., Семенов А.В. Возрастная динамика частоты спонтанных и индуцированных in vitro хромосомных аберраций в лимфоцитах крови человека при естественном и лучевом старении // Радиац. биол. Радиоэкология. 2010. Т. 50. № 3. С. 253–258.
Sigurdson A.J., Ha M., Hauptmann M. et al. International study of factors afecting human chromosome translocations // Mutat. Res. 2008. Vol. 652. № 2. P. 112-121.
Whitehouse C.A., Edwards A.A., Tawn E.J. et al. Translocation yields in peripheral blood lymphocytes from control populations // Int. J. Radiat. Biol. 2005. Vol. 81, № 2. P. 139-145.
Pouzolet F., Roch‑Lefevre S., Giraudet Al. et al. Monitoring translocation by M‑FISH and three‑color FISH painting techniqus: a study of two radiotherapy patients // J. Radiat. Res. 2007. Vol. 48. № 5. P. 425-434.
Suto Y., Akiyama M., Noda T., Hirai M. Construction of a cytogenetic dose-response curve for low‑dose range gamma‑irradiation in human peripheral blood lymphocytes using three‑color FISH // Mutat. Res. 2015. Vol. 794. P. 32-38.
Anderson R.M., Tsepenko V.V., Gasteva G.N. et al. mFISH analysis reveals complexity of chromosome aberrations in individuals occupationally exposed to internal plutonium: a pilot study to assess the relevance of complex aberrations as biomarkers of exposure to high‑LET α particles // Radiat. Res. 2005. Vol. 163. № 1. P. 26-35.
Hande M.P., Azizova TV., Burak L.E. et al. Complex chromosome aberrations persist in individuals many years afer occupational exposure to densely ionizing radiation: an mFISH study // Genes, Chromosomes, Cancer. 2005. Vol. 44. № 1. P. 1-9.
Sotnik N.V., Osovets S.V, Scherthan H., Azizova TV mFISH analysis of chromosome aberrations in workers occupationally exposed to mixed radiation // Radiat. Environ. Biophys. 2014. Vol. 53. № 2. P. 347-354.
Tawn E.J., Whitehouse C.A., Holdsworth D., De Ruyck K., Vandenbulcke K., Tierens H. mBAND analysis of chromosome aberrations in lymphocytes exposed in vitro to α‑particles and γ‑rays // Int. J. Radiat. Biol. 2008. Vol. 84. № 1. P. 1-7.
Сотник Н.В., Азизова ТВ., Осовец СВ. Структурные повреждения
генома у работников плутониевого производства // Радиац. биол. Радиоэкология. 2011. Т. 51. № 2. С. 213-217.
Сотник Н.В., Азизова ТВ. Использование методов mFISH и mBAND в биоиндикации воздействия внутреннего α‑излучения // Радиац. биол. Радиоэкология. 2016. Т. 56. № 2. С. 156-162.
Bedford J.S., Goodhead D.T Breakage of human interphase chromosomes by alpha particles and X‑rays // Int. J. Radiat. Biol. 1989. Vol. 55. № 2. P. 211-216.
Hittelman W.N., Rao P.N. Premature chromosome condensation. I. Visualization of X‑ray induced chromosome damage in interphase cells // Mutat. Res. 1974. Vol. 23. № 2. P. 251-258.
Karachristou I., Karakosta M., Pantelias A. et al. Triage biodosimetry using centromeric/telomeric PNA probes and Giemsa staining to score dicentrics or excess fragments in non‑stimulated lymphocyte prematurely condensed chromosomes // Mutat. Res. 2015. Vol. 793. P. 107-114.
Prasanna P.G., Escalada N.D., Blakely W.F. Induction of premature chromosome condensation by a phosphatase inhibitor and a protein kinase in unstimulated human peripheral blood lymphocytes: a simple and rapid technique to study chromosome aberrations using specifc whole‑chromosome DNA hybridization probes for biological dosimetry // Mutat. Res. 2000. Vol. 466. № 2. P. 131-141.
Durante M., Furusawa Y., Gotoh E. Technical report. A sample method for simultaneous interphase‑metaphase chromosomal analysis in biodosimetry // Int. J. Radiat. Biol. 1998. Vol. 74. № 4. P. 457–462.
Gotoh E., Kawata T., Durante M. Chromatid break rejoining and exchange aberration formation following γ‑ray exposure: analysis in G2 human fbroblasts by chemically in‑ duced premature chromosome condensation // Int. J. Radiat. Biol. 1999 Vol. 75. № 9. P. 1129–1135.
Grossi G., Durante M., Gialanella G. et al. Chromosomal aberrations induced by high‑energy iron ions with shielding // Adv. Space Res.. 2004. Vol. 34. № 6. P. 1358–1361.
Hatzi V.I., Terzoudi G.I., Paraskevopoulou Ch. et al. Te use of premature chromosome condensation to study the ifuence of environmental factors on human genetic material in interphase cells // Sci. World J. 2006. Vol. 6. P. 1174–1190.
Kanda R., Eguchi‑Kasai K., Hayata I. Phosphatase inhibitors and premature chromosome condensation in human peripheral lymphocytes at diferent cell‑cycle phases // Somatic Cell and Molecular Genetics. 1999. Vol. 25. № 1. P. 1–6.
Kanda R., Hayata I., Lloyd D.C. Easy biodosimetry for high‑dose radiation exposures using drug‑induced, prematurely condensed chromosomes // Int. J. Radiat. Biol. 1999. Vol. 75. № 4. P. 441–446.
Kawata T., Ito H., Uno T. et al. G2 chromatig damage and repair kinetics in normal human fbroblast cells exposed to low‑ or high‑LET radiation // Cytogenet. Genome Res. 2004. Vol. 104. № 1–4. P. 211–215.
Lamadrid A.I. Garcia O., Delbos M. et al. PCC‑ring induction in human lymphocytes exposed to gamma and neutron irradiation // J. Radiat. Res. 2007. Vol. 48. № 1. P. 1–6.
Lee R., Yamada S., Yamamoto N. et al. Chromosomal aberrations in lymphocytes of lung cancer patients treated with carbon ions // J. Radiat. Res. 2004. Vol. 45. № 2. P. 195–199.
Romero I., Garcia O., Lamadrid A.I. et al. Assessment of simulated high‑dose partial‑body irradiation by PCC‑R assay // J. Radiat. Res. 2013. Vol. 54. № 5. P. 863–871.
Gonzalez J.E., Romero I., Gregoire E. et al. Biodosimetry estimation using the ratio of the longest:shorter length in the premature chromosome condensation (PCC) method applying autocapture and automatic image analysis // J. Radiat. Res. 2014. Vol. 55. № 5. P. 862–865.
M’kacher R., Maalouf E.E.L., Terzoudi G. et al. Detection and automated scoring of dicentric chromosomes in nonstimulated lymphocyte prematurely condensed chromosomes afer telomere and centromer staining // Int. J. Radiation Oncol. Biol. Phys. 2015. Vol. 91. № 3. P. 640–649.